 La inmunología es un amplio campo que abarca tanto
la investigación básica como las aplicaciones clínicas que tienen que ver con
los mecanismos de defensa del huésped en contra de entidades extrañas (no
propias), conocidas como antígenos, con el reconocimiento de dichas moléculas
antigénicas y con las funciones de defensa del huésped, mediadas por la
inmunidad celular, particularmente aquellas relacionadas con la inmunidad a la
enfermedad, la hipersensibilidad, la autoinmunidad, la inmunodeficiencia, y los
trasplantes. (Jawetz, 2005)
La inmunología es un amplio campo que abarca tanto
la investigación básica como las aplicaciones clínicas que tienen que ver con
los mecanismos de defensa del huésped en contra de entidades extrañas (no
propias), conocidas como antígenos, con el reconocimiento de dichas moléculas
antigénicas y con las funciones de defensa del huésped, mediadas por la
inmunidad celular, particularmente aquellas relacionadas con la inmunidad a la
enfermedad, la hipersensibilidad, la autoinmunidad, la inmunodeficiencia, y los
trasplantes. (Jawetz, 2005)
Inmunidad
 Conjunto de mecanismos fisiológicos que permiten al
animal reconocer una sustancia como extraña y neutralizarla, eliminarla o
metabolizarla, sin dañar los tejidos propios. (Carmona, 1997)
Conjunto de mecanismos fisiológicos que permiten al
animal reconocer una sustancia como extraña y neutralizarla, eliminarla o
metabolizarla, sin dañar los tejidos propios. (Carmona, 1997)
Ramas
de la inmunología
Inmunopatología:
Estudio de las reacciones mórbidas provocadas
por la aparición de un antígeno en el organismo y por la formación consecutiva del anticuerpo correspondiente (conflicto antígeno-anticuerpo). (Medicopedia, 2014)
Inmunocitoquímica o inmunoquímica: técnicas
para localizar proteínas concretas en células,
utilizan anticuerpos específicos marcados
enzimáticamente que dan lugar a un
producto de reacción coloreado. (Lawrence, 2003)
Inmunodiagnóstico:
utilización de anticuerpos para detectar
e identificar, sustancias y agentes patógenos,
por ejemplo virus, bacterias, hormonas, drogas. (Lawrence, 2003)
Inmunogenética:
estudio de la expresión genética de los elementos constituyentes del sistema
inmune y de las enfermedades autoinmunitarias (Anaya, 2005)
Serología:
estudio de los sueros inmunes y de la utilización
de antisueros para caracterizar patógenos, antígenos o células. (Lawrence,
2003)
Tipos de inmunidad
Inmunidad innata o natural: Lawrence, (2003), afirma que esta inmunidad es “la protección
contra infecciones como consecuencia de la activación de mecanismos de defensa
fijados, relativamente inespecíficos” y Roitt (2008) “sistemas antimicrobianos
relativamente inespecíficos que son innatos en el sentido que no son afectados
en forma intrínseca por contacto previo al agente infeccioso”. La inmunidad
natural viene dada desde el nacimiento, y se activa ante cualquier estímulo
nocivo; además, actúa siempre de la misma manera.
la inmunidad innata, como
el término lo sugiere, viene incorporada y se da de una forma “espontánea”,
desde el momento en que se nace, no obstante, existen diferencias entre
poblaciones en la forma en que actúa dicha inmunidad y en la eficiencia de la
misma en función de ciertos patógenos.
Según la raza o etnia del
individuo, pueden existir resistencia o susceptibilidad a ciertas enfermedades
infecciosas. Por ejemplo, existe una mayor resistencia a la malaria entre
poblaciones de raza negra que en las de raza blanca, y los blancos son más
susceptibles a la fiebre amarilla. Asimismo, las especies difieren en cuanto a
la susceptibilidad a infecciones; por ejemplo, el VIH no afecta a los
chimpancés, pero sí a los humanos. Entre individuos existen también
diferencias, como se ve en infecciones que infectan a una cantidad determinada
de personas, pero no todas enferman, e
incluso en casos de epidemias graves, donde parte de la población enferma
muere, pero otra parte no, debido a una mejor respuesta inmunitaria.
Inmunidad adquirida
Es el tipo de inmunidad a una infección producida
por la exposición previa a un patógeno o por la inmunización contra él. (Lawrence,
2003)
Inmunidad
pasiva: tipo de inmunidad a corto plazo
adquirida por transferencia de anticuerpos ya formados, como la que ocurre por
la transferencia de anticuerpos de la madre al feto a través de la placenta y a
través de la leche materna al lactante. (Lawrence, 2003)
Inmunidad
activa: forma de inmunidad adquirida tras la
inmunización a un antígeno de un agente infeccioso. Incluye una primera
etapa donde ocurre la enfermedad y se forman anticuerpos después del primer
contacto con el antígeno, y una segunda donde ocurre el contacto contra el
mismo antígeno pero no se presenta la enfermedad debido a la respuesta de
memoria que se produce, por el primer contacto. (Lawrence, 2003)
Inmunidad natural: tipo de inmunidad que se da de forma espontánea,
es decir, viene “insertada” en el organismo desde el nacimiento. (Lawrence,
2003)
Inmunidad artificial: ocurre cuando se inmuniza a una persona o animal
con anticuerpos (suero) o microorganismos modificados por el hombre en forma de
vacunas. (Lawrence, 2003)
Inflamación
 Tortora (2006) dice que la inflamación es una
respuesta defensiva, inespecífica del organismo ante una lesión tisular.
Asimismo detalla que las causas pueden ser contactos con gérmenes patógenos,
sustancias químicas irritantes, abrasiones, temperaturas extremas y trastornos
o deformaciones celulares. Los signos y síntomas característicos de la
inflamación son rubor, calor, tumefacción (edema) y dolor. Cabe añadir que la
inflamación puede producir falta de función en el área afectada en función del
sitio y extensión de la lesión. El cuerpo, a través de la inflamación, intenta
eliminar a los microorganismos, toxinas o sustancias extrañas presentes en el
sitio de lesión, impedir la diseminación a otros tejidos y preparar al sitio de
lesión para el proceso de reparación tisular en un intento de restaurar la
homeostasis del tejido afectado. La inflamación consiste en A) vasodilatación e
incremento, de la permeabilidad vascular. B) migración de los fagocitos
presentes en la circulación y el líquido intersticial, y C) reparación tisular
Tortora (2006) dice que la inflamación es una
respuesta defensiva, inespecífica del organismo ante una lesión tisular.
Asimismo detalla que las causas pueden ser contactos con gérmenes patógenos,
sustancias químicas irritantes, abrasiones, temperaturas extremas y trastornos
o deformaciones celulares. Los signos y síntomas característicos de la
inflamación son rubor, calor, tumefacción (edema) y dolor. Cabe añadir que la
inflamación puede producir falta de función en el área afectada en función del
sitio y extensión de la lesión. El cuerpo, a través de la inflamación, intenta
eliminar a los microorganismos, toxinas o sustancias extrañas presentes en el
sitio de lesión, impedir la diseminación a otros tejidos y preparar al sitio de
lesión para el proceso de reparación tisular en un intento de restaurar la
homeostasis del tejido afectado. La inflamación consiste en A) vasodilatación e
incremento, de la permeabilidad vascular. B) migración de los fagocitos
presentes en la circulación y el líquido intersticial, y C) reparación tisular
Fiebre
 La fiebre es “la temperatura corporal anormalmente
elevada, producto de ls reprogramación del termostato hipotalámico” (Tortora,
2006). Por lo general, tiene lugar durante procesos infecciosos e
inflamatorios. Muchas toxinas provenientes de las bacterias tienen la capacidad
de elevar la temperatura corporal, y en ocasiones provocan la liberación de
citocinas productoras de fiebre como la Interleucina-1 de los macrófagos. La
elevación de la temperatura corporal potencia la acción de los interferones, inhibe
el crecimiento de algunos microbios e incrementa la velocidad de las reacciones
que contribuyen a la reparación de los tejidos.
La fiebre es “la temperatura corporal anormalmente
elevada, producto de ls reprogramación del termostato hipotalámico” (Tortora,
2006). Por lo general, tiene lugar durante procesos infecciosos e
inflamatorios. Muchas toxinas provenientes de las bacterias tienen la capacidad
de elevar la temperatura corporal, y en ocasiones provocan la liberación de
citocinas productoras de fiebre como la Interleucina-1 de los macrófagos. La
elevación de la temperatura corporal potencia la acción de los interferones, inhibe
el crecimiento de algunos microbios e incrementa la velocidad de las reacciones
que contribuyen a la reparación de los tejidos.
Anticuerpos
Carmona (1997) habla de los anticuerpos y los define
como “sustancias elaboradas por el organismo en respuesta al estímulo provocado
por un antígeno, con el cual se combinan específicamente”.
Los anticuerpos son producidos por las células
plasmáticas, estás células provienen de células B, que son células linfáticas
que al interactuar de formas complejas con linfocitos T de ayuda, se convierten
en células productoras de anticuerpos (células plasmáticas). Los anticuerpos pertenecen a un grupo de proteínas llamadas
globulinas y también se les conoce como inmunoglobulinas. La mayoría de los
anticuerpos posee cuatro cadenas polipeptídicas, dos, idénticas entre sí,
llamadas cadenas pesadas y otras dos llamadas cadenas livianas; las primeras
contienen unos 450 aminoácidos y las segundas unos 220. Las cadenas pesadas se
unen a las livianas por puentes disulfuro y otro puente disulfuro mantiene las
cadenas pesadas unidas. Los extremos de las cadenas pesadas y livianas constituyen los sitios de
reconocimiento de los antígenos, con los cuales se unen en esas zonas. (Tortora, 2006)
Las inmunoglobulinas son 5, y se designan con las
letras G, A, M, D y E. Se representan con la anteposición de las letras Ig a
una de las antes mencionadas; así que las inmunoglobulinas se representan como
IgA, IgM, IgG, IgD y IgE. Las propiedades de las Ig varían, pero pueden
mencionarse: el peso molecular va de 150 mil a 900 mil daltons; solo la IgG se
transfiere a través de la placenta al feto, la IgG y la IgM intervienen en la
vía clásica del complemento, mientras las demás en la vía alterna. Además la Ig
de mayor concentración en suero es la IgG con 75% y la de menor concentración
es la IgE con 0.004 %. (Carmona, 1997)
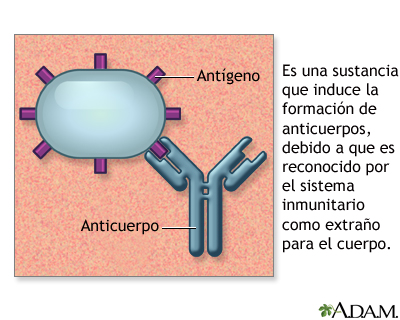
Antígenos
Los antígenos son sustancias que, introducidas en un
organismo animal, da lugar a la formación de anticuerpos, es decir, de células
capaces de reaccionar contra él. De esta definición se desprende que los
antígenos poseen inmunogenicidad, es
decir la capacidad de originar una respuesta inmunitaria y antigenicidad, esto es, la capacidad de reaccionar específicamente.
Por lo común, los antígenos más potentes son de naturaleza proteica, aunque
también los lípidos, los polisacáridos e incluso los ácidos nucleicos, pueden,
en circunstancias determinadas, comportarse como antígenos. Sin embargo, las
células del sistema inmunitario reconocen solo una parte del agente que ingresa
al organismo, es decir, una parte de una molécula que puede generar una
respuesta inmune; a esta parte, que conforma el antígeno, se le denomina
epítopo, o determinante antigénico (Carmona, 1997)
Requerimientos
de la inmunogenicidad
Según Carmona (1997), para que una sustancia sea
inmunógena se necesitan 3 condiciones:
1. Que la sustancia sea extraña: Por
ejemplo, la inyección de albúmina bovina en un conejo, genera respuesta
inmunitaria. Cuanto más extraña sea la sustancia, mayor será la probabilidad de
que sea rechazada.
2. Que sea accesible al sistema
inmunitario: Por ejemplo, existen agentes fúngicos
q no cumplen esta propiedad, como algunos hongos dermatofitos que pueden vivir
muchos años en la piel sin generar respuesta inmunitaria
3. Que sea compatible con la
constitución genética del animal: La respuesta
inmunitaria está sujeta a control genético
y existen variaciones individuales para responder a un antígeno en
particular. Por ejemplo, algunos antígenos son inmunógenos para el ser humano
pero no para el cobayo, así como incluso dentro de una misma especie, algunos
individuos responden a algunos antígenos, y otros no.
Consecuencias
de la unión del anticuerpo con el antígeno
Después de unirse con un antígeno, un anticuerpo
puede producir variados efectos: A) neutraliza o bloquea algunas toxinas
bacterianas y evita la adhesión de
algunos virus a la célula diana. B) Inmovilización bacteriana, debido a que los
anticuerpos se generan por la presencia de antígenos presentes en los cilios o
flagelos de las bacterias móviles, la reacción antígeno-anticuerpo puede causar
la perdida de motilidad de la bacteria, lo cual limita su diseminación a otros
tejidos. C) Aglutinación y precipitación antígenos: esto ocurre, dado que los
anticuerpos presentan dos o más sitios de unión para los antígenos, lo que trae
como consecuencia que se realice una reacción antígeno-anticuerpo cruzada entre
distintos patógenos produciendo la aglutinación (agrupación) de éstos. Las
células fagocíticas digieren a los microorganismos aglutinados con mayor
rapidez. De la misma manera los antígenos solubles secretados pueden precipitar
al reaccionar en forma cruzada con los anticuerpos, y son más fácilmente
fagocitados. D) activación del complemento a través de la puesta en marcha por
parte de los complejos antígeno-anticuerpo de la vía clásica del sistema del
complemento. E) facilitación de la fagocitosis, puesto que el eje de los
anticuerpos sirve como una “bandera” que atrae a los fagocitos, una vez que los
antígenos se unen a los anticuerpos; estos facilitan la actividad de los
fagocitos debido a que producen aglutinación y precipitación de los antígenos,
activación del sistema de complemento y a través de la cobertura de los
microorganismos con anticuerpos (opsonización), lo que los hace más
susceptibles a la fagocitosis. (Tortora, 2006)
Mecanismo
de unión entre anticuerpo
La reacción de unión entre un antígeno y un
anticuerpo depende de una unión química (enlace), en que las moléculas se
mantienen adheridas por fuerzas de atracción. Las moléculas del antígeno tienen
diversos puntos superficiales, de contacto, por lo regular de 10 a 50 puntos
por molécula, y que sirven de lugar de anclaje para el anticuerpo. Éste posee, por lo común, dos puntos que le
permiten actuar como un puente intermolecular, para así integrar una red de
pequeños precipitados o grumos. El sitio de contacto del antígeno es una
pequeña proyección que encaja con unas concavidades en la superficie del
anticuerpo, que produce una conexión que se mantiene por fuerzas
intermoleculares como las fuerzas de Van der Waals, puentes de hidrógeno,
fuerzas hidrófobas y electrostáticas. (Carmona, 1997)
Métodos
para la detección de las reacciones antígeno-anticuerpo
Precipitación
y aglutinación
Los anticuerpos del suero presentan dos tipos de
acciones sobre los antígenos según sea la naturaleza de estos. Si los antígenos
son organismos celulares, los anticuerpos producen aglutinación y ocasionan precipitación
si son moléculas o partículas en solución coloidal. Los anticuerpos reciben el
nombre de aglutininas o precipitinas respectivamente. En general, la
precipitación y aglutinación son muy similares, aunque las diferencias dependen
de la naturaleza del antígeno. (Carmona, 1997)
Mecanismo
y técnica de la precipitación
La unión de un precipitógeno (antígeno) con una
precipitina ocurre en segundos, pero puede tardar de horas a días en ocurrir la
precipitación. El antígeno debe ser multivalente, es decir ha de contar con
varios sitios que le permita combinarse con el anticuerpo, que es divalente, lo
que conlleva a la formación gradual de una red que precipita al alcanzar
ciertas dimensiones. El pH y la temperatura influyen también en el proceso.
La proporción entre antígeno y anticuerpos desempeña
un papel importante en el mecanismo de reacción, lo que se demuestra
experimentalmente al mezclar en tubos de ensayo, cantidades crecientes de
precipitógeno y cantidades constantes de la precipitina. En los tubos que
contienen mayor cantidad de anticuerpos, no ocurre precipitación; en los tubos
donde el precipitógeno va en aumento, ocurre la precipitación y en los que hay
una cantidad mucho mayor de precipitógeno, tampoco ocurre la precipitación,
debido a que no hay suficiencia de anticuerpos.
En la práctica se usa suero puro o de baja dilución
y antígeno en diluciones variables. Pueden mezclarse ambos componentes en un
tubo de ensayo, en cuyo caso, los agregados aparecerán dispersos en el medio.
En la zona de precipitación de observa un anillo en los puntos de contacto.
Esta prueba se denomina reacción del anillo y en el momento en que termina la
precipitación corresponde al título o cantidad de anticuerpos capaces de
reaccionar con la más alta dilución del antígeno. (Carmona, 1997)
Mecanismo
y técnica de la aglutinación
El mecanismo es esencialmente el mismo que el da la
precipitación, a diferencia que el antígeno se ubica sobre una célula o partícula. En este caso los antígenos se llaman
aglutinógenos y los anticuerpos aglutininas. Los antígenos no necesariamente
tienen que estar vivos, porque si están muertos también provocan la reacción.
Si se prepara una suspensión de agentes infecciosos,
en solución salina al 0,9% y se mezcla con el suero correspondiente, se aprecia
como la turbiedad de la suspensión desaparece al formarse los aglutinados que
se depositan paulatinamente en el fondo de un tubo de ensayo. Normalmente las
bacterias poseen una misma carga eléctrica negativa, que las mantiene en
dispersión en los medios que se mantienen suspendidas. Ahora bien, si lacarga
se modifica con la adición de ciertas sales, las bacterias no se repelen, y
caen en el fondo del tubo contentivo de la dilución.
En la aglutinación y en la precipitación se cumplen
dos fases en el mecanismo de reacción. La primera está constituida por la unión
específica antígeno-anticuerpo, y la segunda es llevada a cabo por electrolito,
que modifica el potencial eléctrico superficial, con la consecutiva formación
de los aglutinados. Aún así, cabe recalcar en que los anticuerpos deben vencer
las fuerzas intercelulares de rechazo, derivadas de las cargas negativas de la
superficie celular y de la nube iónica que las rodea.
Asimismo, cabe destacar que, a pesar de estar en
combinación bivalente con el antígeno (glóbulos rojos), algunas aglutininas no
son capaces de producir aglutinación si no se sitúan en un medio rico en
albúmina sérica, ya que de esta forma se vencen las fuerzas electrostáticas de
repulsión de los glóbulos rojo, lo que favorece la proximidad entre ellos, y
por tanto, la aglutinación.
La técnica puede hacerse de forma macroscópica y
microscópica. En la primera, llamada “método macroscópico en tubo”, se deposita
en varios tubos la misma cantidad de microorganismos en solución salina, y se
agrega igual volumen del suero aglutinante, pero en diluciones distintas. Se
mantiene en baño de María, a 37 grados Celsius durante una hora o a 56 ºC
durante 30 minutos; después se observa
la dilución máxima del suero donde hubo aglutinación. Esta dilución del límite
reaccionante representa el título de anticuerpos
En la segunda técnica, llamada “método
microscópico”, se usa una gota de solución del suero y una de antígeno, tomada
con una pipeta de Pasteur de punta fina. Las gotas se colocan en un
cubreobjetos para observarlas al microscopio, según la técnica de gota
pendiente. Es fácil ver como los microorganismos móviles pierden su movilidad y
luego al igual que los inmóviles se adhieren entre sí hasta quedar todos
agrupados en grumos. (Carmona, 1997)
Componentes
físicos del sistema inmunitario
El sistema inmunitario tiene tres funciones
principales:
1. Drenaje del exceso del líquido
intersticial: los vasos linfáticos drenan el exceso
de líquido intersticial desde los espacios tisulares y lo devuelven a la sangre
2. Transporte de los lípidos de la
dieta: los vasos linfáticos se encargan del transporte de
lípidos y vitaminas liposolubles (A, D, E, K) que se absorben desde el tracto
gastrointestinal hacia el torrente sanguíneo
3. Generación de la respuesta
inmunitaria: el tejido inmunológico inicia la
respuesta de elevada especificidad dirigida contra algún microorganismo en
particular o célula anormal. Los linfocitos T y B, junto a los macrófagos
reconocen a las sustancias extrañas y emplean estrategias para atacarlas. (Tortora,
2006)
Capilares
linfáticos
Tienen un
diámetro similar al de los capilares sanguíneos y presentan una estructura única que le permite al líquido intersticial
ingresar en ellos pero no retornar al espacio intersticial. Esto se debe a la
superposición de las células endoteliales de las paredes del capilar. Cuando la
presión del líquido intersticial supera al de la linfa, las células se separan
levemente, como la apertura de una puerta vaivén de un solo sentido y permite
que el líquido intersticial entre en los capilares. Cuando la presión de la linfa aumenta (dentro
de los capilares) respecto a la presión del líquido intersticial, las células
endoteliales se adhieren fuertemente entre sí y evitan la salida de la linfa;
sin embargo, la presión de la linfa disminuye a medida que ésta fluye por los
capilares linfáticos lo que permite el continuo ingreso del líquido
intersticial por diferencias de presión. Junto a los capilares linfáticos hay
filamentos de fijación compuestos por fibras elásticas que se extienden por
fuera de las células endoteliales linfáticas y mantienen a los capilares unidos
a los tejidos circundantes. Por ejemplo, cuando
se acumula líquido intersticial y ocurre edema tisular, los filamentos son
traccionados, lo que ocasiona un aumento de la brecha de las células epiteliales
del capilar, y por consiguiente, un aumento del ingreso del líquido
intersticial al capilar, lo que redunda en una disminución del edema. (Tortora,
2006)
Troncos
y conductos linfáticos
Los vasos linfáticos son de mayor grosor que los
capilares linfáticos y son los encargados de llevar la linfa a los ganglios
linfáticos. Cuando los vasos linfáticos salen del ganglio, se reúnen y forman
troncos linfáticos. Los troncos principales son el lumbar, intestinal, broncomediastínico,
subclavio y yugular. El lumbar recolecta
linfa que proviene de miembros inferiores, las paredes y órganos de la pelvis,
los riñones, glándulas suprarrenales y la pared abdominal. Los troncos
intestinales se encargan de la linfa que viene del estómago, intestinos,
páncreas y parte del hígado. El broncomediastínico recoge linfa del corazón,
pared torácica y pulmones. El tronco subclavio, linfa de los miembros
superiores, y el tronco yugular de la proveniente de la cabeza y cuello.
Los troncos linfáticos drenan la linfa en dos
conductos principales: el conducto torácico y el conducto linfático derecho. El
primero mide entre 38 y 45 cm de longitud y es el principal conducto a través
del cual la linfa retorna a la sangre y recibe linfa de los troncos lumbares
derecho e izquierdo, troncos intestinales, del yugular broncomediastínico y
subclavio izquierdos; por ende, conducto torácico lleva linfa de la porción
izquierda de la cabeza, cuello y tórax, miembro superior izquierdo y la región
inferior a las costillas. El conducto linfático derecho mide aproximadamente
1,2 cm, y recibe la linfa de los troncos yugular, subclavio y
broncomediastínico derechos; así, recoge la linfa de la parte derecha del
cuerpo. Desde el conducto linfático
derecho, la linfa llega a la sangre venosa en la unión entre las venas yugular
interna derecha y subclavia derecha. (Tortora, 2006)
Ganglios
linfáticos
 Tienen forma de alubia y están diseminados por todo
el cuerpo, miden entre 1 y 25 mm de longitud y presentan una cápsula de tejido
conectivo que se extiende al interior del ganglio en forma de trabéculas. La región funcional,
el parénquima se divide en corteza y médula, ubicadas de forma superficial y
profunda respectivamente. En la corteza hayu dos zonas, la corteza externa
tiene agregados ovoides de linfocitos B llamados “nódulos linfáticos” que
cuando están formados principalmente por linfocitos B, son llamados “nódulos linfáticos primarios”. Un nódulo
linfático secundario tiene linfocitos B de memoria y células plasmáticas y
aparece por causa de la estimulación antigénica. Una vez que un linfocito B
reconoce a un antígeno en un nódulo linfático primario, éste evoluciona hacia
uno secundario. Esto ocurre como
resultado del “encuentro antigénico”. La
zona central de un nódulo linfático secundario consiste en una región clara de
células llamada centro germinal en el
cual hay células B, células foliculares dendríticas y macrófagos. Cuando las
células foliculares dendríticas “presentan “ el antígeno, las células B
proliferan y se diferencian a células plasmáticas productoras de anticuerpos o
a células B de memoria, lo cual ayuda con la defensa inmunitaria inmediata y la
que ocurre como respuesta a un ataque del mismo agente.
Tienen forma de alubia y están diseminados por todo
el cuerpo, miden entre 1 y 25 mm de longitud y presentan una cápsula de tejido
conectivo que se extiende al interior del ganglio en forma de trabéculas. La región funcional,
el parénquima se divide en corteza y médula, ubicadas de forma superficial y
profunda respectivamente. En la corteza hayu dos zonas, la corteza externa
tiene agregados ovoides de linfocitos B llamados “nódulos linfáticos” que
cuando están formados principalmente por linfocitos B, son llamados “nódulos linfáticos primarios”. Un nódulo
linfático secundario tiene linfocitos B de memoria y células plasmáticas y
aparece por causa de la estimulación antigénica. Una vez que un linfocito B
reconoce a un antígeno en un nódulo linfático primario, éste evoluciona hacia
uno secundario. Esto ocurre como
resultado del “encuentro antigénico”. La
zona central de un nódulo linfático secundario consiste en una región clara de
células llamada centro germinal en el
cual hay células B, células foliculares dendríticas y macrófagos. Cuando las
células foliculares dendríticas “presentan “ el antígeno, las células B
proliferan y se diferencian a células plasmáticas productoras de anticuerpos o
a células B de memoria, lo cual ayuda con la defensa inmunitaria inmediata y la
que ocurre como respuesta a un ataque del mismo agente.
La zona de la corteza interna del ganglio no
contiene nódulos linfáticos y está formada principalmente por células T y
células dendríticas que arribaron desde otros tejidos. Las células dendríticas
presentan antígenos a las células T, lo que induce su proliferación, así, los
linfocitos T salen del ganglio linfático y se dirigen a las regiones del
organismo con actividad antigénica.
La médula del ganglio contiene linfocitos B, células
plasmáticas productoras de anticuerpos provenientes de la corteza y macrófagos,
todos embebidos en una red de fibras y células reticulares. (Tortora, 2006)
Timo
Es un órgano bilobulado, cuyos lóbulos se mantienen
juntos gracias a una capa de tejido conectivo, y a la vez están separados por
una cápsula de mayor grosor, también de tejido conectivo. Desde la cápsula se
extienden proyecciones llamadas trabéculas, que penetran en cada lóbulo. (Tortora, 2006)
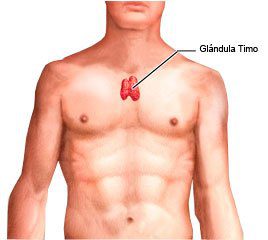 El timo se divide anatómicamente, en primer lugar,
en una corteza donde hay linfocitos T, células dendríticas, células epiteliales
y macrófagos en forma dispersa. Las células pre-T (linfocitos T inmaduros)
migran a la corteza tímica desde la médula ósea roja, donde proliferan e
inician el proceso de maduración. Las células dendríticas, contribuyen al
proceso de maduración. Las células epiteliales de la corteza tienen
prolongaciones que albergan hasta 50 células T, las células epiteliales
colaboran con la selección de células T funcionales en un proceso llamado
“selección positiva”. También producen hormonas para la maduración de las
células T. Los macrófagos ayudan a limpiar los restos de células muertas o
células que están en el proceso de muerte. Las células T que sobreviven
ingresan a la médula.
El timo se divide anatómicamente, en primer lugar,
en una corteza donde hay linfocitos T, células dendríticas, células epiteliales
y macrófagos en forma dispersa. Las células pre-T (linfocitos T inmaduros)
migran a la corteza tímica desde la médula ósea roja, donde proliferan e
inician el proceso de maduración. Las células dendríticas, contribuyen al
proceso de maduración. Las células epiteliales de la corteza tienen
prolongaciones que albergan hasta 50 células T, las células epiteliales
colaboran con la selección de células T funcionales en un proceso llamado
“selección positiva”. También producen hormonas para la maduración de las
células T. Los macrófagos ayudan a limpiar los restos de células muertas o
células que están en el proceso de muerte. Las células T que sobreviven
ingresan a la médula.
En segundo lugar, el timo se divide en una médula,
que tiene linfocitos T maduros, células epiteliales, dendríticas y macrófagos
dispersos en una gran superficie. Algunas de las células epiteliales se agrupan
y forman capas concéntricas de células planas que degeneran y se transforman en
células cargadas de gránulos queratohialinos y queratina, llamadas “corpúsculos
tímicos”. Aunque poco se sabe de ellas,
pueden servir como sitios de muerte de células T presentes en la médula. Las
células T que salen del timo, migran a través de la sangre, hacia bazo,
ganglios linfáticos y otros tejidos linfáticos.
El timo se atrofia con la edad; así, el timo de un
lactante puede pesar 70 g, y en un aciano puede llegar a pesar 3 g. El tejido
del timo se reemplaza con tejido adiposo y areolar. (Tortora, 2006)
Bazo
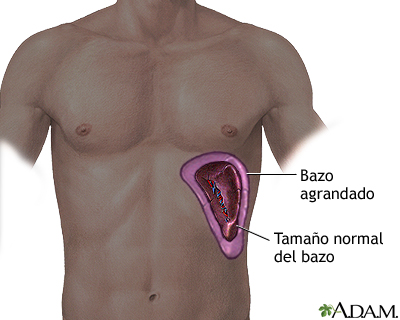 El bazo es un órgano ovoide de aproximadamente 12 cm
de longitud y es la masa de tejido linfático de mayor tamaño. Al igual que el
timo y los ganglios linfáticos, presenta una cápsula y trabéculas; lo que forma
su estroma (estructura de sostén). La parte funcional (parénquima) consta de
pulpa blanca y pulpa roja, la primera es tejido linfático con preponderancia de
linfocitos y macrófagos dispuestos alrededor de las ramas de la arteria
esplénica. En la pulpa roja se encuentran sinusoides venosos cargados de sangre
y cordones tejido esplénico (del bazo), que están formados por glóbulos rojos,
macrófagos, linfocitos, células plasmáticas y granulocitos. Las venas se
encuentran en estrecha asociación con la pulpa roja. (Tortora, 2006)
El bazo es un órgano ovoide de aproximadamente 12 cm
de longitud y es la masa de tejido linfático de mayor tamaño. Al igual que el
timo y los ganglios linfáticos, presenta una cápsula y trabéculas; lo que forma
su estroma (estructura de sostén). La parte funcional (parénquima) consta de
pulpa blanca y pulpa roja, la primera es tejido linfático con preponderancia de
linfocitos y macrófagos dispuestos alrededor de las ramas de la arteria
esplénica. En la pulpa roja se encuentran sinusoides venosos cargados de sangre
y cordones tejido esplénico (del bazo), que están formados por glóbulos rojos,
macrófagos, linfocitos, células plasmáticas y granulocitos. Las venas se
encuentran en estrecha asociación con la pulpa roja. (Tortora, 2006)
Linfocitos
T
 Son glóbulos blancos llamados agranulocitos, puesto
que en el microscopio óptico no son visibles sus gránulos intracelulares,
debido a su pequeño tamaño y su poca capacidad de teñirse. Los hay pequeños y
grandes; el tamaño de los primeros oscila entre 7 y 9 micrómetros de diámetro y
los grandes entre 10 y 14. El núcleo se aprecia redondeado o levemente hendido
y en el citoplasma se forma un halo alrededor del núcleo que se ve
celeste-azulado. Además el citoplasma se hace más visible en tanto la célula
sea más grande. (Tortora, 2006)
Son glóbulos blancos llamados agranulocitos, puesto
que en el microscopio óptico no son visibles sus gránulos intracelulares,
debido a su pequeño tamaño y su poca capacidad de teñirse. Los hay pequeños y
grandes; el tamaño de los primeros oscila entre 7 y 9 micrómetros de diámetro y
los grandes entre 10 y 14. El núcleo se aprecia redondeado o levemente hendido
y en el citoplasma se forma un halo alrededor del núcleo que se ve
celeste-azulado. Además el citoplasma se hace más visible en tanto la célula
sea más grande. (Tortora, 2006)
Activación,
proliferación y maduración de los linfocitos T
Los linfocitos se activan cuando antígenos ingresan
al organismo, y algunos linfocitos que tienen los receptores de membrana
adecuados, pueden reconocer y ligarse a tales antígenos. Estos antígenos ya han
sido procesados con anterioridad y unidos a moléculas del Complejo Mayor de
Histocompatibilidad (CMH) y forman un complejo antígeno-CMH; este “trabajo” lo
realizan las Células Presentadoras de Antígeno (CPA). Las proteínas de membrana
de los linfocitos T llamadas CD4 y CD8 interactúan con el CMH y ayudan a
mantener la unión entre los receptores de membrana de los linfocitos y el CMH,
por lo que se denominan “correceptores”.
Todo el proceso de reconocimiento descrito se
denomina “primera señal de activación de células T”. Sin embargo, se necesita
una segunda señal que en los que intervienen moléculas llamadas
“coestimuladores” y existen una gran variedad de ellas; la interleucina-2 es un
coestimulador. La parte de un coestimulador está en la membrana del linfocito T
y la otra en la de la CPA, y cuando se unen estas células, la unión entre ellas
se prolonga debido a la intervención del coestimulador de las membranas
celulares. Después de ocurridas las dos señales necesarias, se activa el
linfocito T; la activación implica el crecimiento de la célula, su proliferación
y posterior diferenciación, que tiene como resultado la formación de una
población de células T llamada “clon”; tal población celular está ahora
especializada para reconocer el antígeno que activó en primer lugar al
linfocito T; antes de la activación solo unos pocos linfocitos T podían
reconocer al antígeno, pero después existen muchos más que pueden reconocerlo,
lo que mejora de forma significativa la respuesta inmune. La activación,
proliferación y maduración de células T tiene lugar en los tejidos y órganos
linfáticos secundarios. (Tortora, 2006)
Transfusiones
y reacciones contra eritrocitos
La sangre es el tejido que más fácil se puede
trasplantar. Sin embargo, los componentes normales de los glóbulos rojos (GR)
de una persona pueden desencadenar una respuesta antígeno-anticuerpo dañina
para el receptor. En una transfusión incompatible, los anticuerpos del plasma
del receptor se combinan con los antígenos de los GR del donador, lo que
produce aglutinación de los GR. La aglutinación es la respuesta
antígeno-anticuerpo en la cual los GR se entrelazan unos con otros. Cuando se
forman los complejos antígeno-anticuerpo, se activan proteínas plasmáticas de
la familia del complemento. Éstas hacen que las membranas de los GR donados se
vuelva permeable, lo que causa hemólisis de los GR y liberación de hemoglobina
al plasma sanguíneo, lo que puede resultar en insuficiencia renal por bloqueo
de las membranas de filtración. (Tortora, 2006)
Complejo
mayor de histocompatibilidad (CMH)
La capacidad del organismo de reconocer lo propio de
lo ajeno depende en buena medida de un grupo de proteínas de superficie celular
conocidos como “antígenos CMH”. Estos antígenos con codificados por un grupo de
genes denominados Complejo Mayor de Histocompatibilidad (CMH). Existen muchas
formas (alelos) de los genes CMH en los humanos, (unas 40), por lo que existen muchas combinaciones
posibles de los mismos; en consecuencia, es muy improbable que dos personas
tengan los mismos genes CMH a menos que sean gemelos idénticos. Sin embargo,
mientras más relacionados estén los individuos, más similitud hay en sus genes
CMH; dicho sea de paso, existen tres grupos de genes CMH que codifican
distintos conjuntos de proteínas. Los antígenos CMH clase I se encuentran en la
mayor parte de las células nucleadas y son importantes para diferenciar entre
lo propio y lo ajeno. Se unen a antígenos extraños producidos dentro de las
células (por ejemplo, producidos por virus o por injerto de tejido ajeno) y
forman complejos moleculares que se exhiben en la superficie celular. Estos
complejos antígeno-CMH son reconocidos por los linfocitos T citotóxicos. (Solomon,
2001)
Los antígenos CMH clase II se ubican principalmente
en células del sistema inmunitario, en particular en linfocitos B, macrófagos,
algunos linfocitos T y células dendríticas. Tales antígenos regulan las
interacciones entre linfocitos T, linfocitos B y células presentadoras de
antígeno (CPA) y se unen a fragmentos
peptídicos de proteínas que han entrado en la célula a partir de fuentes extrañas,
como bacterias, y se han degradado; luego el complejo antígeno-CMH es exhibido
en la superficie celular y estimula a los linfocitos T de ayuda. Las proteínas
CMH clase III, incluyen componentes del sistema del complemento. (Solomon,
2001)
Presentación
antigénica o de antígenos
Cuando una célula del sistema inmunitario, como un
macrófago, engulle un patógeno, éste es degradado junto con sus antígenos
proteicos. Éstos se degradan en péptidos de menos tamaño, los cuales son unidos
a moléculas del CMH; el complejo resultante se inserta en la membrana
plasmática del macrófago, en un proceso denominado “presentación antigénica”;
una vez inserto en la membrana plasmática celular del macrófago, el complejo
puede ser “presentado” a otras células como los linfocitos T, para su
reconocimiento. Según Tortora (2006) el procesamiento y presentación de
antígenos ocurre de dos maneras:
1.
Procesamiento
de antígenos exógenos: los antígenos externos que ingresaron
al organismo y se encuentran en líquido extracelular, se denominan antígenos exógenos. Estos comprenden, bacterias,
parásitos, polvo, polen y virus que no han infectado aún a las células. Las
Células Presentadoras de Antígenos (CPA) procesan y presentan a los antígenos
exógenos, las CPA son las células dendríticas, macrófagos y células B y se
ubican, de forma estratégica, en los sitios donde es más probable que los
antígenos vulneren los mecanismos de defensa inespecífica, lo que permite su
ingreso al organismo, como la epidermis y dermis de la piel y las mucosas de
los aparatos urinario, reproductor, digestivo, respiratorio y ganglios
linfáticos. El proceso completo se lleva a cabo en distintos pasos:
a.
Internalización
del antígeno: en casi cualquier lugar del cuerpo
donde los “intrusos” lograron burlar las defensas, las CPA internalizan los
antígenos mediante fagocitosis o endocitosis.
b.
Digestión
de los antígenos en fragmentos peptídicos: en el interior
del fagosoma o endosoma, las enzimas digestivas rompen las grandes proteínas
antigénicas y forman pequeños fragmentos peptídicos.
c.
Síntesis
de moléculas del CMH clase II: al mismo tiempo las
CPA sintetizan moléculas CMH-II y las empaquetan en vesículas
d.
Fusión
de vesículas: las vesículas que contienen los
fragmentos de péptidos antigénicos y las que contienen las moléculas CMH-II se
unen y fusionan
e.
Unión
de los fragmentos peptídicos a las moléculas del CMH-II:
Luego de la fusión de ambos tipos de vesículas, los fragmentos de los péptidos
antigénicos se asocian a las moléculas del CMH-II.
f.
Inserción
del complejo antígeno-CMH-II en l membrana plasmática:
la vesícula que contiene los complejos antígeno-CMH-II sufre exocitosis. Como
resultado, dichos complejos se insertan en la membrana plasmática.
2.
Procesamiento
de antígenos endógenos: los antígenos endógenos son las
moléculas antigénicas extrañas que se sintetizan en el interior de las células
del cuerpo. Éstas pueden ser proteínas virales generadas una vez que los virus
infectan a las células utilizando la maquinaria metabólica de la célula huésped
o proteínas anormales sintetizadas por células tumorales. Los fragmentos de los
antígenos endógenos se asocian a moléculas del CMH, en el interior de la célula
infectada, lo que resulta en complejos “fragmento antigénico endógeno-CMH-I”,
que se movilizan hacia la membrana plasmática, donde se expresan en la
superficie celular. El complejo señaliza la presencia de células infectadas que
necesitan ayuda.
Sistema
del complemento
Tortora (2006) habla
del sistema del complemento como “un
sistema defensivo compuesto por 30 proteínas que se sintetizan en el hígado y
que circulan constantemente por el plasma sanguíneo y tejidos del organismo”.
Dichas proteínas destruyen a los microorganismos activando fagocitosis,
citólisis e inflamación.
La mayoría de las
proteínas del sistema del complemento se designan con la letra mayúscula C
desde C1 hasta C9. Normalmente se encuentran inactivas, pero por división
enzimática se subdividen en dos porciones, que son las formas activas de la
proteína. Tales partes se representan con la letra a y b, por ejemplo C3a y C3b
son las formas funcionales de la proteína C3. Otras proteínas del complemento
son los factores B, D y P. Las proteínas del complemento actúan en forma de
cascada, es decir, una reacción desencadena a otra que a su vez, produce la
reacción siguiente. Con cada reacción se generan cada vez más productos, con lo
cual el efecto se amplifica. La activación ocurre de tres formas distintas,
aunque todas tienen como propósito activar a C3. La activación de C3 y las
consiguientes reacciones ocurren así:
a. El
C3 inactivo se divide y forma los fragmentos C3a y C3b
b. El
fragmento C3b se adhiere a la superficie de los organismos, mientras que los
receptores de los fagocitos se unen al mismo C3b. Así, el C3B facilita la
fagocitosis, cubriendo a los microorganismos, proceso que se conoce como
opsonización. La opsonización promueve la adhesión de los fagocitos a los
microbios
c. El
fragmento C3b también inicia una serie de reacciones que provocan citólisis.
Primero C2b divide a C5. El fragmento C5b se une entonces a C6 y C7 los cuales
se unen a la membrana plasmática del microorganismo invasor. Acto seguido, C8 y
varias moléculas de C9 se unen a las demás proteínas del complemento, las
cuales en conjunto forman un complejo d
ataque de membrana, en forma de cilindro, que se inserta en la membrana
plasmática.
d. C3a
y C5a se unen a los mastocitos, y provocan la liberación de histamina por parte
de éstos, lo cual genera aumento de la permeabilidad vascular durante la
inflamación. C5a también atrae a los fagocitos hacia el sitio de inflamación
(quimiotáxis).
La
proteína C3 puede activarse por tres vías. La primera, llamada vía clásica, se inicia cuando los
anticuerpos se unen a los antígenos (microorganismos). El complejo antígeno-anticuerpo formado se
une a C1 y lo activa. Por último, se
activa C3 y el fragmento C3b induce la fagocitosis, citólisis e inflamación. La
segunda vía, se llama vía alterna y no implica la presencia de
anticuerpos. Se inicia por la interacción de complejos de lípidos-hidratos de
carbono sobre la superficie de los microorganismos y los factores del
complemento B, D y P, lo cual activa a C3. La tercera vía, denominada vía de las lectinas, se da porque los
macrófagos que engullen a los microorganismos patógenos, liberan sustancias
químicas que inducen al hígado a producir proteínas denominadas lectinas, que
luego se unen a hidratos de carbono localizados en la superficie microbiana,
que en última instancia activan a C3.
Una
vez que el sistema del complemento se activa, las proteínas plasmáticas y las
proteínas que se encuentran en las células del organismo, como las sanguíneas,
degradan el C3B activado, lo cual disminuye la capacidad destructiva del
sistema del complemento para así minimizar el daño que pueda generarse en los
tejidos propios. (Tortora, 2006)
Inmunoadherencia
Es la fijación en la superficie de glóbulos rojos humanos lavados, no sensibilizados, de microbios que están
aislados o en masas. Este fenómeno se produce en presencia de anticuerpos específicos del microbio y del complemento, y aumenta la fagocitosis de los microbios por los leucocitos. En términos más generales, puede fijar diversos complejos antígenos (soluble
o figurado), anticuerpos, complemento en la superficie de partículas tales como hematíes, plaquetas, granos de sílice o de almidón. (Medicopedia, 2014)
Inmunodeficiencia combinada
La Enfermedad
por Inmunodeficiencia Combinada Grave, que involucra a las células T, B y NK
(células citotóxicas naturales) representa la forma más grave de
inmunodeficiencia primaria y afecta a 1 de cada 80 mil nacimientos vivos. Los
afectados presentan defectos profundos en la inmunidad celular y humoral con
muerte dentro del primer año de vida dado que ocurren infecciones oportnunistas
recurrentes y severas. Son frecuentes las diarreas prolongadas y las neumonías
por Pneumocystis carinii, que es un hongo patógeno, así como candidiasis bucales y dérmicas. Si
se vacunan con organismos atenuados, los afectados fallecen debido a una
infección progresiva. El tratamiento requiere un transplante de médula ósea.
(Roitt, 2008)
Deficiencias de anticuerpos
En ciertas
inmunodeficiencias asociadas a linfocitos B, no se observa reducción del número
de células de este linaje, sino que las células B son incapaces de sintetizar
uno o varios tipos de inmunoglobulinas. El caso más común lo representa la
deficiencia de IgAm, puesto que se da en cada uno de 800 individuos. Los
sintomas más característicos son infecciones sinopulmonares, colitis y
enteritis, es decir, infecciones de mucosas. A menudo, la enfermedad se
encuentra asociada a enfermedades autoinmunes (ejemplo, Lupus) o a enfermedades
alérgicas. Otros individuos presentan una condición denominada hiper-IgM, en el que hay mutaciones que
alteran el diálogo entre células T y B, necesario para el cambio de isotipo, lo
que produce que el linfocito B produzca IgM, pero no IgG, IgA o IgE.
Se han
descrito, asimismo, individuos con deficiencias selectivas en algunos de
isotipos de las IgG. Estos afectados desarrollan con frecuencia infecciones del
tracto respiratorio.
En neonatos,
ocurre una deficiencia de IgG. Las IgG que le traspasa la madre al recién
nacido pueden funcionar de 6 a 12 meses y se solapa con el inicio de la propia
producción de estas inmunoglobulinas por parte del infante. Pero si esta
síntesis se retrasa, ocurre un déficit de inmunoglobulinas, lo que puede acarrear
infecciones. Sin embargo, tal deficiencia es transitoria y desaparece cuando
eventualmente el individuo sintetiza sus propias IgG (Roitt, 2008)
Tipos de inmunidad
Inmunidad activa: ésta de
produce con la administración de una vacuna, que son antígenos hechos
con los agentes causales de las enfermedades o con sus toxinas. Al ser
inyectadas, estimulan la formación de sus respectivos anticuerpos en el
organismo o estimulan una reacción celular defensiva. En general, las vacunas
de agentes vivos atenuados produce un mayor grado de inmunidad que la de
agentes muertos.
Cuando se
inocula por primera vez una vacuna en dosis adecuada, se forman anticuerpos que
aparecen en la sangre después de un período de latencia de 4 a 6 días, y cuya
máxima concentración se alcanza a los 12 o 16 días, luego de lo cual, decrece
hasta un determinado nivel en el que se mantiene por meses.
Sin embargo,
las vacunas operan hasta un límite de eficacia, después del cual, deja de
actuar. Por ejemplo, si pasado un tiempo prudente, se administra una nueva
dosis (segunda dosis), los títulos de anticuerpo se elevarán pronto hasta
superar el nivel alcanzado en el primer estímulo, descienden de forma más lenta
y se mantienen en mayor concentración por un período más prolongado. No
obstante, el incremento de anticuerpos es cada vez menor con cada inyección
adicional de antígeno; por último, se alcanza un título de anticuerpos máximo
que no puede ser superado por inoculaciónes posteriores.
El desarrollo
de la inmunidad es lento. Si un mes más tarde se mide el título de anticuerpos,
se nota que ha bajado en grado apreciable, pero si se aplica una segunda
dosis, menor que la primera, se
incrementan pronto y en un grado notable al término de dos o tres días. La
reacción al estímulo secundario es rápida.
Si transcurre
demasiado tiempo desde la última dosis, se encuentran concentraciones muy bajas
o nulas de anticuerpos, pero el organismo conserva la “experiencia” o “memoria
antigénica” y responderá a la dosis de resfuerzo o de recuerdo en forma rápida
e intensa (reacción amnésica). (Contreras, 1997)
Formas de inmunización activa
a. Inmunización con toxinas y toxoides : El toxoide
se usa hoy en día en lugar de la toxina, por ser más seguro y eficaz. Si se
calienta a 70ºC una toxina tratada con formaldehído pierde su toxicidad, pero
es antigénica (toxoide). Si se añada alumbre al toxoide, su porción antigénica
se precipita y el producto resultante, lavado y resuspendido en solución
salina, tiene la ventaja de provocar un estímulo antigénico prolongado por
cuanto el toxoide se separa poco a poco del alumbre. Si se agrega hidroxido o
fosfato de aluminio, la porción inmunógena se adhiere a las partículas del
compuesto dando lugar al toxoide absorbido. La toxina destoxicada rinde mejores
resultado que la pura y por ello se ha usado en le prevención de
toxoinfecciones como diftería y tétanos.
b. Inmunización con microorganismos muertos: las vacunas
bacterianas se preparan con la siembra de microorganismos de reconocido valor
antigénico en medios sólidos adecuados; luego se suspende el cultivo resultante
en solución salina estéril, de manera que se obtenga una concentración de 100 a
1000 000 000 de bacterias por mililitro. Los agentes son muertos por sustancias
químicas (fenol, formaldehido, etc.) o agentes fídicos como luz ultravioleta y
calor. En el producto final se efectúan
pruebas de esterilidad (no debe contener bacterias vivas) , inocuidad (no ser
patógenos ni provoca reacciones locales o generales) y potencia (eficaz en la
protección). Las vacunas se recomiendan en la profilaxia de enfermedades como
la fiebre tifoidea, tos ferina, cólera, y peste así como en infecciones
piogénicas localizadas y en la defensa orgánicacontra egentes bacterianos de
unfección secundaria. Las vacunas virósicas se preparan con solo inocular
conejos o embriones de pollo y las vacunas antirickettsia se elaboran en huevos
fecundados de pollo.
c. Inmunización con organismos vivos atenuados: se hace en
medios disgenésicos (hace difícil la reproducción) , por pases en animales,
huevos fecundados y medios de cultivo. Por ejemplo, el cultivo del bacilo de la
tuberculosis de origen bovino en medio de bilis proporcioné la cepa denominada
BCG, que constituye el agente inmunizante contra esa enfermedad y cuya acción
no depende de la función de los anticuerpos sino del entrenamiento de las
células monocíticas del sistema defensivo. El pase del virus de la rabia en
conejos, lo transforma de virus de la calle, en virus fijo. El pase del virus
rábico en embriones de pollo provoca su mutación, con cambios en su carácter
patógeno. (Carmona, 1997)
Inmunidad
pasiva
Este tipo de inmunización implica la administración de un
suero que contiene anticuerpos, por tanto, no es necesario que transcurra el
período de latencia que exige la inmunidad activa para formar sustancias
protectoras, sino que tales sustancias se dan al paciente con la debida rapidez
y en la cantidad requerida.
La inmunidad conferida por sueros es transitoria puesto que
los anticuerpos desaparecen a las dos o tres semanas. La protección de suero
homólogo (de la misma especie) va
seguida de una rápida eliminación de un 50% del total administrado, para luego
disminuir lentamente. Cuando el suero es heterólogo, el segundo acontecimiento
ocurre de forma acelerada, quizás debido a la formación de anticuerpos y a la
subsiguiente estructuración de grandes complejos antígeno-anticuerpo, que son
retirados de la circulación por el sistema fagocitario. Existen tres tipos de
suero:
Los sueros
inmunes antitóxicos: son antitoxinas que neutralizan
la toxina y no ejercen acción sobre las bacterias que las producen. Así, la
antitoxina diftérica, neutraliza la toxina en la circulación, pero no ejerce
efecto alguno en el bacilo diftérico que se desarrolla en los tejidos. Entre
los sueros más comunes de este tipo se encuentran: antitetánico, antidiftérico,
antigangrenoso, antibotulínico y antiescarlatínico.
Los sueros
antibacterianos: prácticamente en desuso desde la
llegada de las sulfas y antibióticos, fueron muy empleados para atacar las
infecciones por meningococos, neumococos, estafilococos y bacilo disentérico.
Los sueros
antivirósicos: son muy eficaces, aunque su campo de acción es
limitado en la medicina humana; de ellos, el más conocido es el antirrábico
(Carmona,1997)
No hay comentarios:
Publicar un comentario